|
|
Царство Животные (Zoa), Подцарство Многоклеточные (Metazoa), Надраздел Эуметазои (Eumetazoa), Раздел Двусторонне-симметричные (Bilateria), Тип Членистоногие (Arthropoda), Подтип Хелицеровые (Chelicerata), Класс Паукообразные, или Арахниды (Arachnida), Отряд Пауки (Aranei)
Как говорилось, паутинные бородавки — это видоизменённые брюшные ножки. Они расположены на брюшке снизу перед бугорком с анальным отверстием. На концах бородавок имеются многочисленные хитиновые паутинные трубочки (видоизменённые волоски), которыми открываются протоки паутинных желёз  . У лифистиоморфных пауков паутинных бородавок четыре пары: две пары наружных, развивающихся из зачатков конечностей, и две пары внутренних, образующихся как выросты стенки тела. Бородавки здесь мпогочлениковые, чем напоминают настоящие ножки. У большинства пауков сохраняются три пары бородавок: две пары наружных, обычно 2-члениковых, и пара задних срединных, нечленистых. В ряде семейств и родов пауков наблюдается дальнейшее сокращение числа бородавок до двух и даже до одной пары, но пауки, вовсе лишённые паутинных бородавок, неизвестны. Передняя медиальная пара бородавок у пауков группы Cribellatae превращена в так называемый крибеллум — небольшую пластинку, пронизанную порами, через которые выделяется вещество особых паутинных желёз. У некоторых пауков отдельные пары бородавок удлинены, иногда бывают даже длиннее туловища. . У лифистиоморфных пауков паутинных бородавок четыре пары: две пары наружных, развивающихся из зачатков конечностей, и две пары внутренних, образующихся как выросты стенки тела. Бородавки здесь мпогочлениковые, чем напоминают настоящие ножки. У большинства пауков сохраняются три пары бородавок: две пары наружных, обычно 2-члениковых, и пара задних срединных, нечленистых. В ряде семейств и родов пауков наблюдается дальнейшее сокращение числа бородавок до двух и даже до одной пары, но пауки, вовсе лишённые паутинных бородавок, неизвестны. Передняя медиальная пара бородавок у пауков группы Cribellatae превращена в так называемый крибеллум — небольшую пластинку, пронизанную порами, через которые выделяется вещество особых паутинных желёз. У некоторых пауков отдельные пары бородавок удлинены, иногда бывают даже длиннее туловища.
Паутинные железы помещаются в полости брюшка, в большинстве случаев хорошо развиты и многочисленны. Проток каждой желёзки открывается на конце паутинной трубочки. Наряду с обычными трубочками имеется небольшое число так называемых паутинных конусов, на которых открываются протоки более крупных желёз. Относительно примитивные формы паутинных желёз свойственны лифнстио- и мигаломорфным паукам, у которых имеются пучки мелких овальных желёзок, но уже здесь число их может достигать нескольких сотен. Аранеоморфные пауки обладают паутинными железами нескольких типов, производящими паутину разных сортов. Наиболее сложен паутинный аппарат у пауков-крестовиков (семейство Araneidae), у которых различают до шести типов паутинных желёз, причём на паутинных бородавках в общей сложности имеется более 500 трубочек и около 20 паутинных конусов. Паутина, выделяемая железами разных типов, используется для различных целей. Так, крестовик для построения ловчей сети употребляет секрет всех своих желёз, кроме так называемых трубковидных, паутина которых идёт на яйцевые коконы. Паутинная рама и натягиваемые внутри радиусы делаются из сравнительно толстых сухих нитей, выделяемых ампуловидными железами. При этом несколько желёзок действует одновременно и отдельные тонкие нити соединяются жидким секретом в более толстый «кабель». Так называемые грушевидные железы выделяют пучки тонких волокон, которыми концы основных нитей прикрепляются к окружающим предметам. Спиральная нить делается из паутины дольковидных желёз и покрывается липким секретом древовидных желёз, который не застывает на воздухе и вскоре собирается в микроскопические капельки. Благодаря этому ловчие тенета более или менее длительно остаются липкими. Особенно долго сохраняют клейкость тенета пауков группы Cribellatae. На ситовидной крибеллярной пластинке открываются протоки множества (до 600) особых паутинных желёзок. Выделяемая паутина энергично расчёсывается задними ногами, на предпоследнем членике которых у крибеллятных пауков для этого имеются ряды крепких щетинок (так называемый каламиструм). Получается особого рода «пряжа»; осевые нити окружаются слизистой муфтой, внутри которой имеется нить, изогнутая многочисленными петлями. Из такой слизистой паутины крибеллятные пауки строят ловчие сети разной формы, очень липкие и долго сохраняющиеся. Так называемые грушевидные железы выделяют пучки тонких волокон, которыми концы основных нитей прикрепляются к окружающим предметам. Спиральная нить делается из паутины дольковидных желёз и покрывается липким секретом древовидных желёз, который не застывает на воздухе и вскоре собирается в микроскопические капельки. Благодаря этому ловчие тенета более или менее длительно остаются липкими. Особенно долго сохраняют клейкость тенета пауков группы Cribellatae. На ситовидной крибеллярной пластинке открываются протоки множества (до 600) особых паутинных желёзок. Выделяемая паутина энергично расчёсывается задними ногами, на предпоследнем членике которых у крибеллятных пауков для этого имеются ряды крепких щетинок (так называемый каламиструм). Получается особого рода «пряжа»; осевые нити окружаются слизистой муфтой, внутри которой имеется нить, изогнутая многочисленными петлями. Из такой слизистой паутины крибеллятные пауки строят ловчие сети разной формы, очень липкие и долго сохраняющиеся.
Паутина по химическому составу близка к шёлку шелкопрядов, от которого отличается малым содержанием склеивающего вещества — серицина, растворимого в воде. Основу паутинного шёлка составляет протеид фиброин, образованный сложным комплексом альбуминов, α-аланина и глютаминовой кислоты. По физическим свойствам паутина также близка к гусеничному шёлку, но гораздо эластичнее и прочнее. Нагрузка разрыва для паутины составляет от 40 до 261 кг на 1 мм2 сечения нити (у некоторых Araneus), в то время как для гусеничного ш`лка — только 33—43 кг на 1мм2.
На вентральной стороне брюшка ближе к основанию открываются дыхальца. Дыхательный аппарат пауков интересен тем, что в этом отряде происходит смена лёгких трахеями. Примитивные пауки имеют две пары лёгких. Таковы лифистио- и мигаломорфные пауки (четырёхлёгочные прежней классификации) и небольшое семейство Hypochilidae среди аранеоморфных пауков. У подавляющего большинства последних (двулёгочные прежней классификации) на месте задней пары лёгких развиваются трахеи, а у нескольких семейств (Caponiidae и других) трахеи образуются и на месте передней пары лёгких. Таким образом, среди пауков имеются четырёхлегочиые, дышащие только лёгкими, двулёгочные, дышащие лёгкими и трахеями, и безлёгочные, дышащие только трахеями. Лёгкие пауков по строению сходны с лёгкими других арахнид.
Трахеи отличаются разнообразием строения и у большинства пауков развиты относительно слабо. Различают трахеи местного и общего значения. Первые представлены пучками коротких, обычно неветвящихся трубочек, не выходящих за пределы брюшка. Вторые более длинные, иногда анастомозирующие и ветвящиеся, проникают через стебелёк брюшка в головогрудь и её конечности. У большинства двулёгочных пауков имеются четыре неветвящпхся трахейных ствола. Дальнейшее усложнение трахей проявляется в образовании на их концах многочисленных тонких трубочек, причём стволы средней пары проникают в головогрудь, например, у пауков-скакунов. Наиболее дифференцированы трахеи у безлёгочных пауков. От передних стигм отходят пучки коротких тонких трахей, которые омываются кровью особого трахеального синуса, то есть функционируют как своеобразные вторичные лёгкие. От задних стигм в головогрудь и брюшко отходят толстые стволы, которые ветвятся и доставляют кислород непосредственно к органам и тканям, то есть функционируют, как и положено трахеям. Кутикулярные стенки трахей имеют опорные палочковидные утолщения, очень похожие на палочковый аппарат лёгочных карманов, препятствующий их спадению. Реже трахеи имеют спиральные утолщения, которые так характерны для трахей насекомых, а среди арахнид — сольпуг и сенокосцев.
У большинства пауков, имеющих трахеи, лёгочное дыхание всё же преобладает. В связи с этим хорошо развита кровеносная система. Замазывание стигм лёгких вазелином приводит к быстрому снижению работы сердца и в дальнейшем к смерти. Только у форм с хорошо развитыми трахеями отключение лёгких не вызывает заметных нарушений жизнедеятельности. О пауках в целом можно сказать, что они, заселяя самые различные места обитания, в том числе и сухие, не слишком «спешили» сменить лёгкие трахеями и донесли до нас все стадии этого процесса. В этом усматривается проявление паутинной деятельности пауков, благодаря которой они всюду создают для себя убежище с подходящим микроклиматом.
Центральная нервная система пауков сильно концентрирована. Органы чувств играют важную роль в их сложной жизни. Преобладающее значение, особенно у тенетных форм, имеет осязание. Туловище и придатки покрыты многочисленными осязательными волосками. Особого строения волоски — трихоботрии имеются на педипальпах и ногах. Их бывает до 200. С помощью трихоботрий паук ощущает самые незначительные дуновения воздуха, например, от пролетающей мухи. Трихоботрии воспринимают ритмические колебания в широком диапазоне частот, но не непосредственно как звук, а через вибрацию паутинных нитей, то есть как осязательные ощущения. Если прикоснуться к сети паука-крестовика звучащим камертоном, паук направляется к нему как к добыче. Однако звук камертона, не прикасающегося к сети, обращает паука в бегство. Считают, что звук при этом воспринимается какими-то иными органами. Известно, что пауки нередко выходят на сеть при звуке музыкального инструмента, например, скрипки. При такой положительной реакции, очевидно, имеет место не слуховое, а осязательное ощущение резонирующих нитей паутины.
Другая разновидность осязательного чувства — восприятие степени натяжения паутинных нитей. При изменении их натяжения в опыте паук разыскивает своё убежище, двигаясь всегда вдоль наиболее натянутых нитей. Крестовик гораздо проворнее бежит к тяжёлому предмету, попавшему в сеть, нежели к лёгкому.
Органы равновесия и слуха неизвестны у пауков, но они обладают этими чувствами. Овладев добычей, паук возвращается в центр сети. Если поместить муху в тенета над центром, паук направляется к ней вверх. Поворотом сети на 90 или 180° можно дезориентировать паука. Покончив с мухой, он начинает спускаться по сети, как бы к её центру, и оказывается у края сети. В этом случае чувство тяжести и равновесия преобладает над изменившимися осязательными ощущениями.
Наличие у пауков слуха подтверждается рядом фактов. Пауки-ликозиды реагируют на жужжание спрятанной мухи, которую они не видят, аранеиды поднимают передние ноги на звук определённого тона. Некоторые пауки издают звуки, причём в отдельных случаях доказана их роль в привлечении полов. Звуковые органы представляют собой стридуляторы, то есть трущиеся друг о друга поверхности с рёбрышками или рядами щетинок. Они бывают на хелицерах и педипальпах или только на хелицерах, на соприкасающихся частях головогруди и брюшка и в других местах. Звуковой аппарат имеют либо только самцы, либо оба пола. Последнее наблюдается у части мигаломорфпых пауков, у которых на хелицерах и педипальпах имеются ряды особых щетинок (гребешок и лира). Паук быстро трёт ими друг о друга. Звуки, издаваемые мелкими пауками (семейства Theridiidae, Linyphiidae и другие), очень слабы и регистрируются только специальными приборами. Их высота — 325—425 колебаний в секунду. Некоторые мигаломорфные пауки издают звуки, слышимые человеком,— треск, жужжание, шипение. В ряде случаев звук сочетается с позой угрозы и имеет, по-видимому, предостерегающее значение.
Органами обоняния служат тарзальные органы на лапках передних ног и лировидные органы, имеющиеся в большом числе на туловище и придатках. Пауки различают запахи летучих веществ, но реагируют обычно на близком расстоянии от источника запаха. Самцы по запаху отличают тенета половозрелой самки от тенет неполовозрелой. Роль запаха в данном случае доказана экспериментально. Если из паутины или оторванной ножки половозрелой самки сделать эфирную вытяжку и налить её в блюдечко, то после испарения эфира посаженный в блюдечко самец проявляет характерное половое возбуждение. Тарзальные органы служат и вкусовыми, с их помощью паук в опыте различает чистую воду и растворы различных веществ. По-видимому, эти органы играют роль при отыскании питьевой воды, необходимой некоторым паукам. Чувствительные вкусовые клетки найдены в стенках глотки пауков. В опыте пауки хорошо отличают кусочки сердцевины бузины, пропитапные питательным раствором, от таких же кусочков, но пропитанных водой. Первые высасываются, а вторые удаляются из тенет.
Зрение пауков несовершенно, особенно у тенетных форм. Лучше видят бродячие пауки, особенно активные днём. Глаз обычно четыре пары. Передние медиальные глаза, называемые главными, тёмные; остальные, побочные глаза, обычно блестящие благодаря отражающей свет внутренней оболочке (зеркальцу). Размеры и взаимное положение глаз различны в разных систематических группах пауков 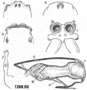 . Чаще они образуют два поперечных ряда, но бывают расположены и иначе. Иногда отдельные пары глаз увеличены, например, четыре передних глаза у пауков-скакунов, медиальные задние глаза у Dinops (семейство Dinopidae). В ряде случаев число глаз сокращается до шести, четырёх или двух. Среди пещерных пауков есть слепые. Глаза тенетных пауков расположены так, что охватывают большое поле зрения, но различают они в основном силу и направление света, улавливая движение крупных объектов. Многие сидящие на тенетах пауки замечают приближающегося человека и падают на паутинной нити. При резком изменении привычного освещения окружающих предметов норковые пауки теряют ориентировку и не могут сразу найти своё логово. Пауки-бокоходы (семейство Thomisidae), подстерегающие добычу иа цветах, замечают бабочку-капустницу на расстоянии 20 см, а муху только на расстоянии 3 см. Бродячие ликозиды имеют обширное поле зрения и видят движущееся небольшое насекомое на расстоянии 20—30 см, но не различают его форму. . Чаще они образуют два поперечных ряда, но бывают расположены и иначе. Иногда отдельные пары глаз увеличены, например, четыре передних глаза у пауков-скакунов, медиальные задние глаза у Dinops (семейство Dinopidae). В ряде случаев число глаз сокращается до шести, четырёх или двух. Среди пещерных пауков есть слепые. Глаза тенетных пауков расположены так, что охватывают большое поле зрения, но различают они в основном силу и направление света, улавливая движение крупных объектов. Многие сидящие на тенетах пауки замечают приближающегося человека и падают на паутинной нити. При резком изменении привычного освещения окружающих предметов норковые пауки теряют ориентировку и не могут сразу найти своё логово. Пауки-бокоходы (семейство Thomisidae), подстерегающие добычу иа цветах, замечают бабочку-капустницу на расстоянии 20 см, а муху только на расстоянии 3 см. Бродячие ликозиды имеют обширное поле зрения и видят движущееся небольшое насекомое на расстоянии 20—30 см, но не различают его форму.
Своего рода исключение представляют небольшие пауки-скакуны (семейство Salticidae). Их длиннофокусные главные глаза дают на сетчатке крупное изображение при малом поле зрения (как в фотокамере с телеобъективом). В отличие от других глаз, зрительные элементы сетчатки здесь густо расположены, благодаря чему зрение предметное; на расстоянии 8 см паук видит муху в деталях. Малое поле зрения этих глаз компенсируется замечательной особенностью: они могут двигаться при помощи специальных мышц. Паук следит глазами за добычей — пример среди наземных членистоногих редкостный. Побочные глаза не различают форму предметов, но зато расположены так, что паук замечает всякое движение впереди, позади себя и над собой. Передние боковые глаза имеют общее бинокулярное поле зрения около 40°, благодаря чему паук воспринимает объёмность предметов н расстояние до них. Глаза скакунов действуют как единый зрительный аппарат. Если муха приближается к пауку сзади, он замечает её движение задними глазами на расстоянии 20—25 см и поворачивается к ней так, что она попадает в поле зрения передних глаз. Теперь она воспринимается отчетливее и в пространстве. Затем паук ловит её главными глазами, воспринимает крупным планом и начинает следить за ней глазами. На расстоянии 8 см объект узнается как добыча, с 4 см паук начинает подкрадываться и с 1,5 см молниеносно прыгает на муху с такой точностью, что редко промахивается. Хорошее зрение скакунов помогает им передвигаться в траве, ловко прыгая с листочка на листочек. С помощью глаз самец обнаруживает самку, а будучи ослеплён, не узнаёт её и не производит своих характерных брачных танцев. Посаженный перед зеркалом, самец скакуна реагирует на своё изображение как на соперника, принимает позу угрозы или бросается на него.
Скакуны и некоторые другие пауки различают цвет предметов. Это установлено несколькими методами, в том числе выработкой условных рефлексов. Паукам предлагались мухи при красном и синем и при красном и зелёном освещении. Красное освещение сопровождалось раздражением электрическим током. После нескольких повторении опыта паук брал муху только при синем или зелёном свете.
Активность пауков зависит от температуры и влажности воздуха, большинство их тепло- и влаголюбиво, но есть немало видов, активных при сравнительно низкой температуре. В зимние оттепели, при +6, +7 °С, некоторые мелкие пауки иногда в массе появляются на снегу. Пауки, в особенности тенетные, подобно многим насекомым, чутко реагируют и на изменения барометрического давления, благодаря чему известны как «предсказатели погоды». Действительно, они усиленно строят тенета в хорошую погоду, а перед ненастьем прячутся в свои убежища и нередко даже перестают реагировать на добычу, попавшую в тенета.
Пауки очень прожорливые хищники, питаются главным образом насекомыми, которых они высасывают. Способы ловли добычи разнообразны. Бродячие пауки подкарауливают добычу, крадутся к ней и настигают одним или несколькими прыжками. При этом уже у бродячих форм добыча нередко опутывается паутинными нитями. Пауки, ведущие оседлую жизнь, почти всегда делают те или иные ловчие устройства из паутины, от простых сигнальных нитей, протянутых от устья логовища, в котором сидит паук, до очень сложных ловчих сетей.
Убивается добыча, как правило, ядом. Ядовитые железы очень характерны для пауков. У примитивных форм железы небольшие, целиком помещаются в хелицерах, для Araneomorphae характерны крупные железы, вдающиеся в полость головогруди. Каждая из двух желёз окружена спиральной мускулатурой, при сокращении которой яд через отверстие на конце когтевидного членика хелицер впрыскивается в тело жертвы. На небольших насекомых яд действует почти мгновенно, но более крупные некоторое время продолжают биться в сетях. Добыча опутывается паутиной, но, если она крупна и паук не может с ней справиться, он сам освобождает добычу. Нередко из тенет выбрасываются и насекомые с резким запахом — клопы и другие.
Объекты питания очень разнообразны. Обычно это самые различные насекомые, подходящие по размеру. В сеть тенетных пауков попадают главным образом летающие формы, чаще других двукрылые. Паук, высасывающий муху,— картина для каждого само собой разумеющаяся. Видовой состав потребляемых насекомых зависит от местообитания и времени года. Многие пауки, живущие близ почвы или в норках, питаются в основном жуками и прямокрылыми. Некоторые наряду с насекомыми затаскивают в норку и поедают дождевых червей и улиток. Есть пауки, специализировавшиеся в выборе добычи, охотящиеся только на муравьев или только на пауков других видов (семейство Mimetidae). Водяной паук Argyroneta охотится за личинками водных насекомых, рачками, мальками рыб. Громадные тропические пауки-птицеяды при случае убивают и съедают небольших птиц, хотя в основном питаются насекомыми. В неволе птицеяды охотно поедают небольших ящериц, лягушек, мышей. Бразильский птицеяд Grammostola питается молодыми змеями и уничтожает их в большом количестве. Амфибиотические пауки (Dolomedes и другие), бегающие по воде, вылавливают небольших рыбок, головастиков и личинок насекомых.
По способу питания пауки в наиболее законченной форме представляют арахнидный тип. Фильтрующий аппарат предротовой полости и глотки, узкий пищевод, мощный сосательный желудок — все это приспособления для питания жидкой пищей. Поймав и убив добычу, паук разрывает и разминает её хелицерами, изливая при этом пищеварительный сок, растворяющий внутренние ткани. Выступающая жидкость всасывается. Выделение сока и всасывание капель пищи чередуются, паук поворачивает жертву, обрабатывая её с разных сторон, пока не останется сморщенная шкурка. Пауки, питающиеся насекомыми с твёрдым покровом, например, жуками, наносят хелицерами рану в сочленовную перепонку, чаще между головой и грудью. Через это отверстие проникает пищеварительный сок и высасывается содержимое жертвы.
В пищеварении и экскреции у пауков существенна роль крупной печени, в клетках которой происходит внутриклеточное переваривание пищи и всасывание. Часть клеток печени, перегруженных экскретом, выходит в просвет кишечника и смешивается в клоаке с белыми выделениями мальпигиевых сосудов. Экскременты выбрасываются мелкими каплями перед новым приёмом пищи.
Часть 1
Часть 2
Часть 3
Часть 4
Часть 5
Тип Членистоногие
Тип Членистоногие (окончание)
Подтип Хелицеровые
Подтип Жабродышащие
Подтип Трилобиты
Подтип Трахейнодышащие
Подтип неопределён
В начало страницы
|
|